Cellular aspects of reproduction and development
Gametogenesis
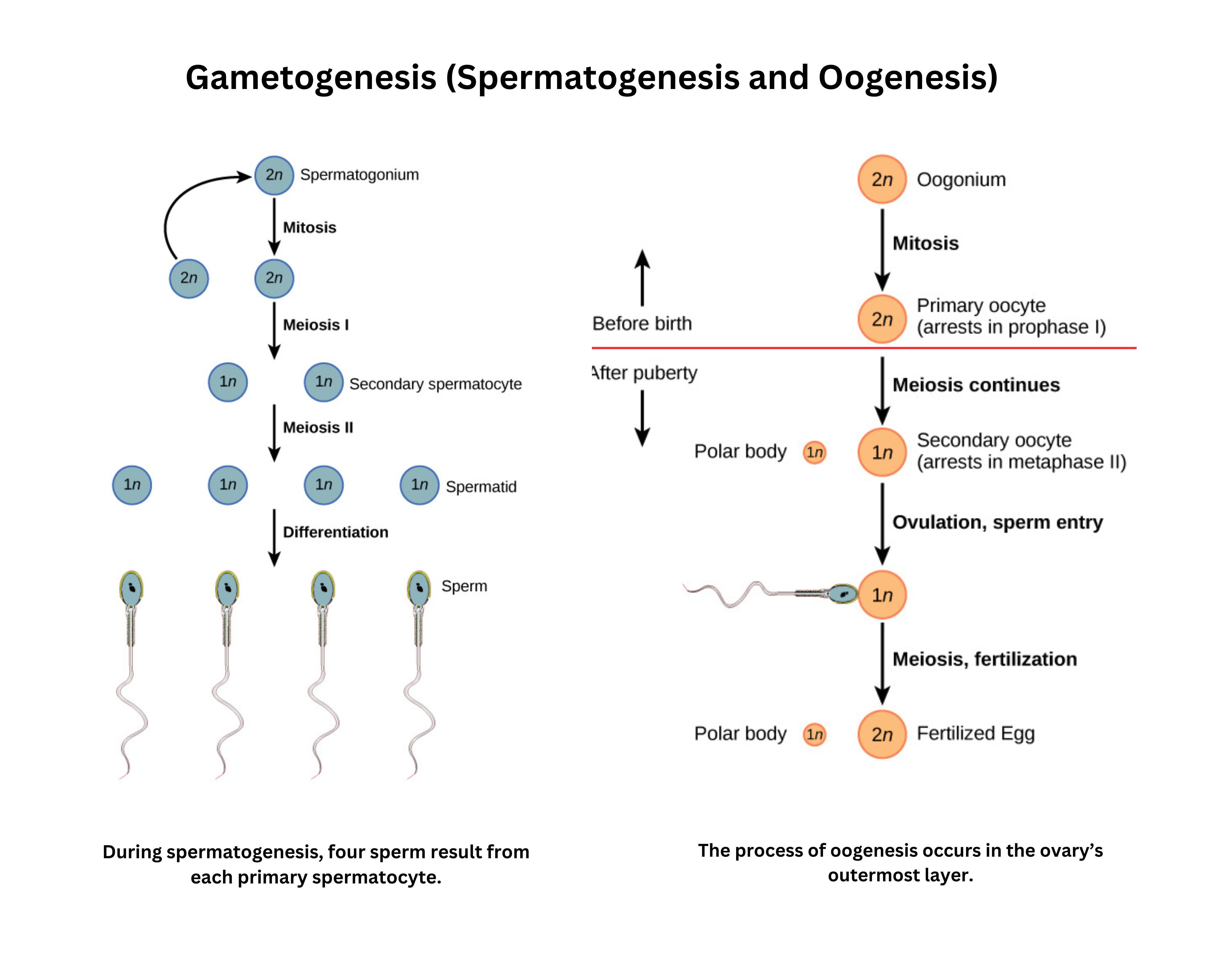
Gametogenesis by meiosis produces haploid cells (gametes). The details differ in males and females, but the overall goal is the same: reduce the chromosome number from diploid (2n) to haploid (n).
In males, spermatogenesis takes place in the seminiferous tubules of the testes:
- A spermatogonium (2n) either self-renews or, after puberty, undergoes mitosis to form a primary spermatocyte (2n).
- The primary spermatocyte enters meiosis I to produce two secondary spermatocytes (n). Each secondary spermatocyte then undergoes meiosis II to form spermatids (n).
- These immature cells mature into motile sperm, also known as spermatozoa.
In females, oogenesis occurs in the ovaries and subsequently in the fallopian tubes:
- An oogonium (2n) replicates by mitosis to yield a primary oocyte (2n), which is arrested in prophase I before birth.
- Each month from puberty to menopause, one primary oocyte resumes meiosis I to generate a secondary oocyte (n), which then pauses at metaphase II.
- Fertilization is required to complete meiosis II, producing the functional ovum (n).
Male and female gametogenesis differ significantly
- Males continually renew spermatogonia and produce four sperm for each primary spermatocyte, whereas females stop renewing oogonia before birth, and each primary oocyte yields a single ovum and polar bodies.
The sperm contributes mainly DNA upon fertilization, while the egg (or ovum) contributes DNA plus mitochondria and other organelles.
Reproductive sequence
- Fertilization:
- Occurs when sperm and egg unite to form a zygote, initiating the reproductive sequence that continues with implantation and embryonic development.
- Implantation
- The zygote develops into a morula (a solid ball of cells), then a blastocyst (in mammals) that implants in the endometrium.
- Development
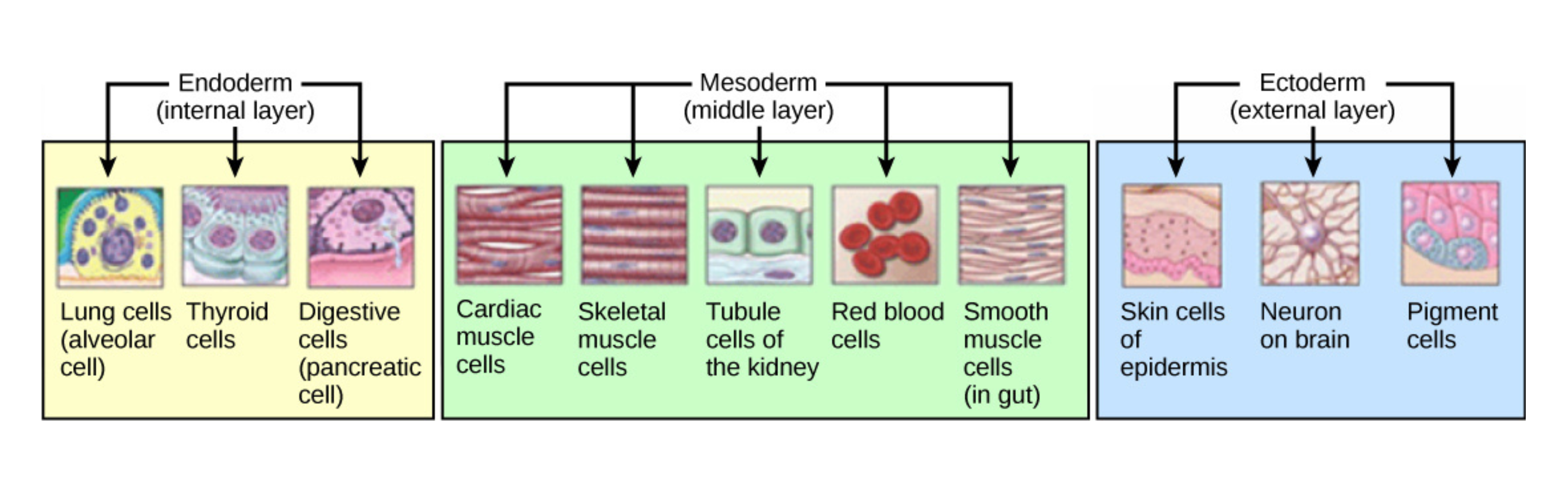
- Early embryogenesis proceeds through gastrulation, during which cells migrate to form three germ layers - endoderm, mesoderm, and ectoderm - followed by organogenesis (e.g., neurulation for the brain and spinal cord).
- Birth
- Ultimately requires a shift in nutrient and oxygen supply from the maternal system to the infant’s independent circulation and respiration.
Embryogenesis
Embryogenesis begins at fertilization. The sperm’s acrosomal reaction and the egg’s cortical reaction help ensure that only one sperm enters.
After fertilization, cleavage divides the zygote into smaller, undifferentiated cells called blastomeres. During cleavage, the total size of the embryo does not increase; instead, the original cytoplasm is partitioned into more cells.
At the blastocyst stage, implantation occurs. Next comes gastrulation, a key stage marked by the first major cell movements and the formation of tissue layers that later give rise to organ systems:
- The endoderm (gut structures)
- Mesoderm (muscles, bones, kidneys, and gonads)
- Ectoderm (skin and nervous system).
Neurulation
- Next, part of the ectoderm becomes neuroectoderm, which will form the nervous system.
- Neuroectodermal cells form the neural plate by differentiating into neuroepithelium.
- As these cells change shape, the tissue folds inward to create a neural groove along the dorsal side, bordered by neural folds.
- When the neural folds converge, they form the neural tube beneath the ectoderm.
- Cells that detach from the neural folds become the neural crest. These cells migrate away from the emerging central nervous system (CNS) and contribute to the formation of the peripheral nervous system (PNS).
- As development continues, each germ layer differentiates into distinct tissues and organs.
Transition to independent homeostasis at birth in mammals
- In mammals, the reproductive sequence of fertilization, implantation, organogenesis, and birth produces an individual with organ systems capable of independent life.
- After birth, the neonate shifts from maternal oxygen and nutrient support to autonomous breathing and feeding, and fetal circulatory shunts close to establish normal blood flow patterns.
Environment-gene interaction in development
- Gene-environment interaction describes how an individual’s genome and their physical and social surroundings work together to shape the traits they exhibit.
- Genes provide the blueprint for development, while factors such as climate, diet, social relationships, and lifestyle can influence how genetic instructions are carried out.
- These interactions help determine the phenotype, or observable characteristics, of an organism.
- For example, a person might inherit a predisposition for a particular condition, but whether the condition manifests can depend on environmental factors like nutrition, stress, or exposure to pollutants.
Mechanisms of development
Cell specialization
- Developmental mechanisms describe how cells specialize, communicate, migrate, and die to form a functional organism.
- Cell specialization occurs in two major phases: commitment and differentiation.
- Commitment has two steps:
- Specification, which can be reversed
- Determination, an irreversible decision to adopt a particular cell fate
- Once determined, cells differentiate by expressing proteins suited to their function. For example, epidermal cells produce keratin for protection, while myocytes synthesize actin and myosin for contraction.
Different tissues reflect these specialized lineages:
- Epithelial (such as skin or organ linings)
- Connective (including blood, bone, tendons, ligaments, and cartilage)
- Nervous (brain, spinal cord, and peripheral nerves)
- Muscle (skeletal, smooth, and cardiac fibers)
Cell-cell communication in development
- Cell communication in development often occurs through induction, in which an inducer signals a nearby responder cell to change.
- Induction can involve direct physical contact (juxtracrine) or the secretion of signaling molecules (paracrine).
- Examples include induction of the lens by the optic vesicle or the influence of dermal mesenchyme on feather development.
Cell migration and pluripotency
- Development also depends on cell migration, as in gastrulation and neural crest migration.
- When migration fails, developmental disorders can result, such as Hirschsprung’s disease.
- Some cells remain pluripotent (e.g., stem cells in bone marrow) and can differentiate into multiple lineages (such as blood or bone cells).
Gene regulation in development
- Gene regulation coordinates the developmental program.
- At the DNA level, modifications such as methylation can silence or activate genes, while histone modifications change chromatin structure.
- RNA processing, including selective export of RNA transcripts or alternative splicing, also changes gene output.
- Regulation of translation (for example, prolonging mRNA lifespan) affects how much protein is produced.
- Post-translational regulation, including enzymatic activation and ubiquitin-mediated degradation, fine-tunes protein function.
Programmed cell death and potential regeneration
- Programmed cell death, or apoptosis, removes unnecessary or damaged cells.
- During apoptosis, cells activate proteolytic enzymes called caspases that break down cellular components in an orderly way without triggering inflammation.
- Normal developmental examples include removal of tissue between digits and resorption of a tadpole’s tail.
- In some organisms, regenerative capacity is notable. Amphibians like newts can dedifferentiate stump cells to regrow limbs.
Senescence and aging
- Senescence and aging reflect a gradual decline in cellular replication and repair capacity.
- This decline is often linked to telomere shortening and the accumulation of DNA damage over time.
- If this damage is not repaired, cells can enter permanent arrest, contributing to the aging process.